
Early Embryology
Embryogenesis
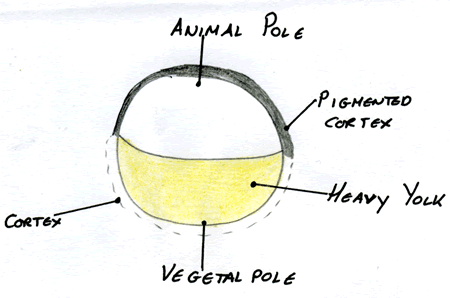
In order to understand how the early brain develops one must have an appreciation of how the embryo is created. Life starts with an egg that has two well definable poles: an animal pole/ epiblast and a vegetal pole/ hypoblast which can be defined by factors such as heavy density substances. The vegetal pole contains the yolk which provides the nutrients required for maintaining cell survival until the formation of extra embryonic tissues (such as the placenta in humans) arise. In the Xenopus egg, once it has been laid by the mother it sinks to the bottom of the pond where heavy cytoplasmic determinants sink to the bottom layer. This leads to differentiation of the bottom layer, as different transcription factors become localised and activated here. In the Xenopus egg the main vegetal transcription factor is VgT; a T-box Transcription factor Once this egg is fertilised by the sperm, it starts to divide into a compact ball of cells called the morula and at some point due to changes in osmolarity, an influx of water forms a central space called the blastocoel. The non VgT expressing pole forms the embryo proper and is divided into 3 germ layers from which all tissue types derive:
1) Ectoderm
2) Mesoderm
3) Endoderm
These form as VgT induces the expression of a member of the TGF-β signalling family Vg1, and these act together to induce the expression of another TGF-β family member called Nodal. It is Nodal that acts to induce the animal tissue closest to it from an ectodermal to a mesodermal or endodermal fate.
| An egg prior to fertilisation showing the pigmented vegetal area, the animal cap, and the cortex that undergoes cortical rotation. Image courtesy of Cassy Ashman |
Dorsal and Ventral Patterning of the Embryo
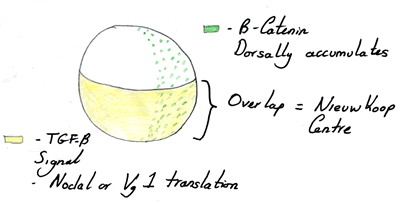
The sperm not only causes cleavage of the egg, but also marks the site of the organiser region of the egg, which occurs opposite to the site of sperm entry. Once the sperm has fertilised the egg the outer cortex moves in a process known as cortical rotation 90° in relation to the rest of the egg. This activates a signalling pathway on one side of the egg called the Wnt signalling pathway and marks the dorsal side of the embryo. It also results in areas of transcription factor expression, with the vegetal hemisphere being divided into a nodal only expressing area and a nodal and Wnt expressing area. The Wnt signalling pathway causes the stabilisation of a signalling factor called β-Catenin which alters the nucleus and leads to an altered gene expression profile. It is the area of overlap between the nodal expressing areas and the β-Catenin expressing areas that causes the formation of the Nieuwkoop centre and thus the organiser region or the Spemann centre.
| An egg showing that overlap of expression to create the organiser region. Image courtesy of Cassy Ashman. |
Gastrulation
The organiser region is an area that is able to undergo self differentiation into what is known as axial mesoderm and begins convergent extension: the formation of a long rod like structure. It is composed of 3 types of progenitor cells: Pharyngeal endoderm, prechordal mesoderm and ectoderm. This rod then migrates inside the cells of the animal hemisphere in a process called gastrulation marking the future anterior/ posterior axis of the embryo in a process that is instrumental in building the head. Once gastrulation is complete the axis of the future embryo are in place.